
Динамічні моделі в біології
Моделі в екології Моделі водних екосистем
Основні особливості водних екосистем
Головний об'єкт досліджень гідроекології - водні екосистеми. Більшість екологів розглядають екосистему як "локалізовану в просторі і динамічну в часі сукупність спільно мешкають і входять до спільноти різних організмів і умов їх існування, що знаходяться в закономірного зв'язку між собою і утворюють систему взаємообумовлених біотичних і абіотичних процесів" (Алімов, 2001). В результаті взаємодії організмів між собою і навколишнім середовищем всередині екосистеми організовуються потоки речовини, енергії та інформації. Екосистема може бути представлена як біорізноманіття видів плюс взаємозв'язок потоків речовини, енергії та інформації, причому останні розглядаються як організують і регулюють (Bayers, Odum, 1993). Динамічна взаємодія потоків речовини, енергії та інформації, що забезпечує стабільність екосистеми в часі в конкретних умовах середовища, і є її функціонування.
Методологічною основою екологічних досліджень служить системний підхід, орієнтований на розкриття цілісності об'єкта і забезпечують її механізмів, на виявлення різноманітних зв'язків складного об'єкта і зведення їх в єдину теоретичну картину. Важливо, щоб в основі екологічних теорій лежали конкретні механізми процесів, а не феноменологічні пояснення або міркування за аналогією (Wiens, 1992).
Вивчаючи різноманітні явища у водних екосистемах, дослідники зустрічаються з різними, в ряді випадків протилежно спрямованими процесами, сукупність і загальний підсумок яких висловлюють у вигляді різного роду балансів. Особливе місце в екосистемі займає баланс органічних речовин, які можуть синтезуватися в самому водоймі і надходити в нього у вигляді зовнішнього навантаження зі стоком річок і при випаданні атмосферних опадів. Він охоплює широке коло різнорідних і складних біотичних процесів у водоймах, з ним пов'язані баланси біогенних і багатьох інших елементів. Балансовий підхід став одним з основних способів кількісних дослідження закономірностей функціонування водних екосистем (Загальні основи вивчення водних екосистем, 1979; Алімов, 1989; Алімов, 2001).
Багато природні системи, по всій видимості, можна розглядати як самоорганізовані критичні системи (Jorgensen et al., 1998; Milne, 1998). Такі системи поєднують в собі концепцію організованої критичності з концепцією самоорганізації, заснованої на розгляді нерівноважних систем і вивченні ієрархії механізмів зворотного зв'язку. Передбачається, що екосистеми функціонують в стані динамічної рівноваги на кордоні хаосу. Швидкості процесів в таких системах збалансовані таким чином, щоб в максимально можливій мірі (оптимально) використовувати доступні ресурси. І в той же час навіть незначний дисбаланс в параметрах і швидкостях може призводити на окремих проміжках часу до хаотичного поведінки і перебудови всієї системи в цілому або окремих її компонентів.
Екологія має справу з принципово нередуціруемого ієрархічними системами (Азовський, 2001; Jorgensen, 1999). Ієрархічність системи виражається в тому, що кожен її елемент (функціональна одиниця) також являє собою систему, а сама вона, в свою чергу, є елементом системи більш високого рівня організації. При цьому такі системи є відкритими, тобто функціональні одиниці як одного, так і різних рівнів пов'язані потоками енергії, речовини та інформації (Азовський, 2001; Алімов, 2001). Таким чином, можна говорити не тільки про ієрархію структур, а й про ієрархію протікають в них процесів (Allen, Starr, 1982; Kolasa, Pickett, 1989; King, 1997).
Починаючи з основоположною роботи Аллена і Старра (Allen, Starr, 1982), концепція ієрархічної організації біологічних систем розвивається в нерозривному зв'язку з проблемою характерних масштабів. Кожен процес має свої характерні масштаби простору і часу, в межах яких формується структура певного рівня. Слід зауважити, що поняття характерних масштабів неоднозначно і по-різному трактується різними авторами. Основна увага при цьому приділяється виділенню явищ, які спостерігаються в істотно різних просторово-часових масштабах, і виявлення відповідних їм структуроутворюючих процесів. Регуляторні механізми в екосистемах функціонують на різних часових масштабах: від декількох хвилин до декількох десятків років. У роботі (Азовський, 2001) проаналізовано зв'язок між характерними масштабами простору 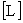 і часу
і часу 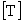 для природних ієрархічних систем різної природи. Показано, що наявні дані добре апроксимуються статечним співвідношенням виду
для природних ієрархічних систем різної природи. Показано, що наявні дані добре апроксимуються статечним співвідношенням виду 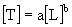 , Де b - параметр масштабу (експонента Херста), a - перехідний коефіцієнт. Характер зв'язку між просторовими і тимчасовими характеристиками, що розвиваються структур в загальному випадку залежить від типу переважаючих структуроутворюючих процесів. Так, якщо поширення структурообразующего процесу в просторі відбувається випадковим (ненаправленим) чином, то характерні розміри утворюється структури збільшуються пропорційно квадратному кореню від часу розвитку. У цьому випадку параметр масштабу дорівнює 2. Класичною моделлю таких систем є випадковий (броунівський) процес поширення речовини або енергії - дифузія (Okubo, 1980). Направлене (в сенсі "захоплення" простору) розвиток процесів призводить до пропорційної зміни просторових і часових масштабів (показник ступеня дорівнює 1). Нарешті, значення експоненти Херста, менші одиниці, показують, що "захоплення" простору структурою відбувається швидше, ніж зростає характерний час її існування. Це свідчить про когерентном (узгодженому в просторі і часі) поведінці окремих елементів такої структури, оскільки процес її формування протікає "майже одночасно" по всьому об'єму простору. Для планктонних екосистем, пов'язаних з водними масами, характерні значення b, які варіюють від 1 (процеси прямого перенесення) до 2 (випадкові дифузійні процеси).
, Де b - параметр масштабу (експонента Херста), a - перехідний коефіцієнт. Характер зв'язку між просторовими і тимчасовими характеристиками, що розвиваються структур в загальному випадку залежить від типу переважаючих структуроутворюючих процесів. Так, якщо поширення структурообразующего процесу в просторі відбувається випадковим (ненаправленим) чином, то характерні розміри утворюється структури збільшуються пропорційно квадратному кореню від часу розвитку. У цьому випадку параметр масштабу дорівнює 2. Класичною моделлю таких систем є випадковий (броунівський) процес поширення речовини або енергії - дифузія (Okubo, 1980). Направлене (в сенсі "захоплення" простору) розвиток процесів призводить до пропорційної зміни просторових і часових масштабів (показник ступеня дорівнює 1). Нарешті, значення експоненти Херста, менші одиниці, показують, що "захоплення" простору структурою відбувається швидше, ніж зростає характерний час її існування. Це свідчить про когерентном (узгодженому в просторі і часі) поведінці окремих елементів такої структури, оскільки процес її формування протікає "майже одночасно" по всьому об'єму простору. Для планктонних екосистем, пов'язаних з водними масами, характерні значення b, які варіюють від 1 (процеси прямого перенесення) до 2 (випадкові дифузійні процеси).
Відповідно до теорії адаптивного циклу (Carpenter et al., 2001), динамічні системи, до числа яких відносяться екосистеми і спільноти, проходять через наступні чотири характерні фази: швидкого зростання і активного використання ресурсів середовища (r - фаза), фазу консервації (стагнації ) (К - фаза), фазу колапсу (руйнування, деструкції) ( 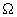 - фаза) і відновлення (реконструкції) (
- фаза) і відновлення (реконструкції) ( 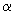 - фаза). Найбільш важливою особливістю цього циклу є існування щодо коротких періодів, протягом яких в системі відбуваються найбільш істотні зміни - фази
- фаза). Найбільш важливою особливістю цього циклу є існування щодо коротких періодів, протягом яких в системі відбуваються найбільш істотні зміни - фази 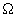 і
і 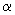 , І більш тривалих періодів еволюційного, поступового, розвитку системи - фази r і K. Це дозволяє провести, хоча б в принципі, поділ змінних на швидкі і повільні (виділити характерні масштаби часів) і спростити тим самим модельне вивчення такого роду систем. В
, І більш тривалих періодів еволюційного, поступового, розвитку системи - фази r і K. Це дозволяє провести, хоча б в принципі, поділ змінних на швидкі і повільні (виділити характерні масштаби часів) і спростити тим самим модельне вивчення такого роду систем. В 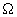 - фазі деякі компоненти і системні атрибути вихідної системи можуть бути втрачені (наприклад, зникнути деякі види). Але потім в наступній,
- фазі деякі компоненти і системні атрибути вихідної системи можуть бути втрачені (наприклад, зникнути деякі види). Але потім в наступній, 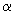 - фазі, в оновленій системі можуть з'явитися нові види, нова система зв'язків і інтеграційних властивостей. Далі, в r - фазі, система переходить на нову траєкторію свого розвитку (з точки зору термінології теорії динамічних систем) і її еволюція має місце в межах цілком певного аттрактора. Протягом довгого, досить повільно розвивається процесу від r до K фазі ймовірність виникнення суттєвих змін в системі значно знижується, хоча вона може стає все більш складною в результаті збільшення кількості внутрішньосистемних зв'язків. На різних етапах адаптивного циклу еластичність системи і її стійкість до зовнішніх впливів можуть бути абсолютно різними. Таким чином, для того щоб ввести міру еластичності і стійкості системи, розробити структуру її математичної моделі, необхідно встановити, на якому масштабі часу розвивається система або на якому масштабі ми хочемо її вивчати. Масштаб, в свою чергу, буде впливати на виділення швидких і повільних змінних, компонентів модельної системи та її параметрів.
- фазі, в оновленій системі можуть з'явитися нові види, нова система зв'язків і інтеграційних властивостей. Далі, в r - фазі, система переходить на нову траєкторію свого розвитку (з точки зору термінології теорії динамічних систем) і її еволюція має місце в межах цілком певного аттрактора. Протягом довгого, досить повільно розвивається процесу від r до K фазі ймовірність виникнення суттєвих змін в системі значно знижується, хоча вона може стає все більш складною в результаті збільшення кількості внутрішньосистемних зв'язків. На різних етапах адаптивного циклу еластичність системи і її стійкість до зовнішніх впливів можуть бути абсолютно різними. Таким чином, для того щоб ввести міру еластичності і стійкості системи, розробити структуру її математичної моделі, необхідно встановити, на якому масштабі часу розвивається система або на якому масштабі ми хочемо її вивчати. Масштаб, в свою чергу, буде впливати на виділення швидких і повільних змінних, компонентів модельної системи та її параметрів.
Багато природних процесів, як вже було зазначено вище, по-різному проявляють себе в залежності від масштабів, на яких вони вивчаються і розглядаються. Екологічні явища принципово є масштабно-залежними як у часі, так і в просторі (Wiens, 1992). Певне уявлення про часових масштабах фізичних і біологічних процесів у водних екосистемах дає наступна таблиця.
Таблиця. Деякі тимчасові масштаби природного мінливості і найбільш важливі масштаби гідробіологічних процесів (Reynolds, 1990).
Фізичні змінні і біологічні процесиТимчасові масштаби (сек) 10-2 10-1 100 101 102 103 104105 106 107 108 109 1010 1011 мікротурбулентність пульсації Мінливість поверхневої освітленості Тривалість часу вітрового перемішування в мілководних озерах Сезонні зміни термічного режиму водойми Межгодовая мінливість гідрофізичних процесів Кліматична мінливість Цикли біогенних елементів Зміна пропорцій між Лімі ірующімі елементами Процеси евтрофікації і перехід екосистеми в новий стан Процеси флуоресценції Швидкість фотосинтезу Швидкість дихання Тривалість розмноження Процеси споживання речовин Процеси асиміляції Формування видової структури співтовариства
Ігнорування цих залежностей може призводити до артефактів при моделюванні. Різке відмінність масштабів часу є внутрішня властивість системи, а не суб'єктивна оцінка її спостерігачем (Молчанов, 1992). У математичної моделі це означає наявність малого параметра в системі, (в найпростіших випадках) можливість такого вибору змінних, яке зводить до розпаду системи на швидку і повільну. Це має першорядне методологічне значення. В результаті ми отримуємо можливість виділити якийсь "квант", структурний елемент і звести задачу до вивчення тільки двох сусідніх масштабів часу. Вивчення взаємодії двох сусідніх за масштабами систем неминуче передбачає повне і послідовне кінетичне розгляд і класифікацію можливих типів поведінки.
При переході від однієї області просторово-часових масштабів до іншої, як правило, відбувається і зміна сукупності факторів, що визначають цілісне поведінку всієї системи або окремих її частин. Такий перехід може бути достатньо різким і на певному часовому проміжку супроводжуватися складними нелінійними ефектами і хаотичним поведінкою, як це нерідко має місце, наприклад, при фазових переходах в фізичних системах. Наші передбачення і пояснення можуть бути досить успішними в межах певних областей просторово-часових масштабів, але абсолютно не працювати в зонах переходів або на інших масштабах. Виділення цих областей і визначення правил переходу між ними є дуже складне завдання, але ці питання ніяк не можна ігнорувати.
Одна і та ж реальна екосистема може моделюватися абсолютно різними моделями при вивченні її на різних масштабах простору і часу. Природні системи являють собою певну ієрархію коливальних систем, "вкладених" один в одного, по крайней мере в сенсі масштабів часу (Молчанов, 1992). При моделюванні таких систем необхідно завжди задаватися питанням, "про які масштабах часу йде мова?", І враховувати тільки ті змінні, періоди яких можна порівняти з досліджуваним масштабом. Повільні змінні можна вважати постійними, а від швидких залишаться тільки середні значення. Статистичні схеми виникають, як правило, при відкиданні швидких змінних. Тому завжди можна говорити прямо про "осреднении" в міру в фазовому просторі, уникаючи терміна "ймовірність" (Молчанов, 1992).
Серед найбільш загальних і важливих особливостей водних екосистем можна відзначити наступні:
· Складність внутрішньої будови;
· Многофакторность впливів зовнішнього середовища;
· Незамкнутість (інформаційна, енергетична, морфологічна, структурна);
· Суттєва нелінійність структурно-функціональних зв'язків;
· Істотна неоднорідність і гетерогенність;
· Висока швидкість біотичного кругообігу;
· Трофічні мережі в багатьох спільнотах включають велику кількість всеїдних організмів;
· За відсутності екстремальних умов чисельність популяцій і співтовариств організмів регулюється або обмеженням ресурсів, або пресом хижаків;
· Структурна і функціональна стабільність екосистем забезпечується за рахунок складних регуляторних процесів, серед яких велике значення мають позитивні і негативні зворотні зв'язки;
· "Буферний" характер реакції природних систем на впливу;
· Контрінтуітівное характер реакції природних систем на зовнішні впливи.
Складність екосистеми формується не тільки великим числом взаємодіючих компонентів, а також прямих і зворотних зв'язків між ними. Ця складність, якщо так можна висловитися, набагато складніша, оскільки екосистеми відносяться до класу складних адаптивних систем. Зворотні зв'язку (їх число і структура) теж безперервно змінюються так, щоб досягти найбільш ефективного (в певному сенсі) використання екосистемою доступного їй ресурсу і забезпечити максимальне пристосування, збереження і виживання входять до неї видів і співтовариств. По суті справи ми ніколи не спостерігаємо екосистему в одному і тому ж стані двічі, а тільки між двома щодо схожими станами. Екосистеми демонструють високу ступінь гетерогенності в часі і просторі і є надзвичайно динамічними системами. Все це і багато іншого якраз і створює ті значні труднощі при вивченні, виділення найбільш істотних властивостей екосистеми і її моделюванні.
З чисто математичної точки зору складність внутрішньої будови екосистеми означає, що її фазовий простір багатовимірний. Біологічні об'єкти складні не тільки внутрішньо. Вони і функціонують в складній, нерідко швидко мінливому середовищі. Більш того, є серйозні підстави думати, що сама складність їх будови носить "компенсаторний" характер. Екосистеми саме тому і складні, щоб у відповідь на будь-які (але не катастрофічні) впливу середовища розвинути внутрішню захисну реакцію таким чином, щоб зберегти максимально незмінною свою внутрішню структуру. Звідси випливає математичне наслідок (Молчанов, 1992): математична модель біологічної системи повинна містити багато параметрів (як безперервних, так і дискретних), задають складну середу, в якій функціонує дана вивчається система.
Оскільки екосистеми водойм є відкритими системами, то звідси випливає важливий висновок про необхідність спільного моделювання як біологічної системи, так і середовища її функціонування. Необхідний синтез гідробіологічних, гідрохімічних і гідрофізичних знань про динаміку екосистеми. Ідея такої єдності усвідомлювалася і висувалася вже з найперших досліджень в області біоокеанологіі і лімнології (Харвей, 1934; Хатчінсон, 1969). К.А Хайлов (Хайлов, 1971), Т. А. Айзатуллін, А.В. Леонов (Айзатуллін, Леонов, 1975; Айзатуллін, Леонов, 1977; Айзатуллін, 1980) вперше у вітчизняній практиці звернули увагу на необхідність адекватної математичної формалізації біогідрохіміческіх процесів, на важливість розгляду циклів з'єднань біогенних елементів (вуглецю, азоту, фосфору, кремнію, кисню) в їх взаємозв'язку, на вивчення механізмів сполучення різних циклів при аналізі закономірностей динаміки біомас гідробіонтів і концентрацій біогенних речовин у водному середовищі, а також запропонували хіміко-кінетичний апарати т вивчення складних біохімічних, хімічних і фізико-хімічних процесів для математичного моделювання біотрансформації речовини в воді. Фундаментальні роботи в цьому напрямку Т.А. Айзатулліна охоплюють практично весь спектр проблемних питань біогідрохіміі, що вивчає хіміко-екологічні, біохімічні та трофодінаміческіе зв'язку.
Одним з найбільш характерних властівостей біологічних об'єктів Взагалі и екосистем зокрема є величезне діапазон (як правило) нелінійніх вплівів, в межах которого система нормально працює. Нелінійність в біології має експонентній характер (Молчанов, 1992). Найбільш яснім вираженість цього є відомій фізіологічній закон "Вебера - Фехнера", Який встановлює логаріфмічну залежність Реакції від впліву. Еволюційній сенс настолько сильною нелінійності Цілком зрозумілій. Логарифмічна шкала відповідей - єдина можливість охопити все інформаційного значущі впливу, зберігши при цьому скільки-небудь прийнятним чином структурні особливості системи.
Екосистеми є істотно неоднорідними і гетерогенними системами (Леонов, 1991; Wiens, 1992). Серед усього різноманіття існуючих в природних умовах форм речовини виділяють відрізняються за своєю природою органогенні елементи (їх називають також біофільние, біогенними або органічними), такі як вуглець, азот, фосфор, сірка, кисень, агреговані фракції яких взаємопов'язані і підрозділяються на зважені і розчинені, біологічні і хімічні. Взаємодії цих фракцій впливають на умови і спрямованість процесів трансформації речовини і конкретних біогенних елементів, а також безпосередньо формують якість водного середовища. Роль зважених і розчинених форм біогенних елементів у функціонуванні водних екосистем виключно велика, хоча в кількісному відношенні ще не достатньо вивчена. До складу зважених компонентів біогенних елементів входять живі організми, діяльність яких безпосередньо визначає розвиток процесів трансформації речовин і круговорот біогенних елементів, а також детрит, який сам відрізняється значною неоднорідністю. Просторова неоднорідність (мозаїчність, плямистість) розподілу планктону має місце в дуже широкому діапазоні масштабів - від десятків і сотень кілометрів в океанах і морях (перші десятки - сотні метрів в озерах і водосховищах) до метрів і навіть десятків сантиметрів. Вона є неодмінною особливістю існування планктонних угруповань. Внаслідок цього розвиток гідробіологічних процесів може визначатися не тільки властивостями займаного ділянки або шару води, але також залежати і від властивостей сусідніх ділянок.
Специфічною особливістю водних екологічних систем є наявність пелагического спільноти. Саме воно має основне значення для біотичного кругообігу речовини у водоймі. Для мілководних, добре перемішуються водойм, велике значення має також і донне співтовариство (Алімов, 2001).
Найважливішим властивість водних екосистем є висока швидкість біотичного кругообігу (Ащепкова, 1978). Цьому сприяють як рухливість водного середовища, добре перемішування, умови, сприятливі для бактеріального розкладання, розчинення і розповсюдження продуктів мінералізації, так і інтенсивний обмін за рахунок малого розміру продуцентів (фітопланктон) і консументів (зоопланктон). Фітопланктон у відкритих водах з'їдається майже відразу після продукування, тобто значна частина продукції проходить через травоїдних. Ці особливості водних екосистем вимагають використання спеціальних математичних моделей.
Трансформація речовини здійснюється у водному середовищі гідробіонтами в процесі споживання органічних і мінеральних компонентів, виділення продуктів обміну і відмирання. Нарощування біомаси гідробіонтів відбувається паралельно з розвитком трансформації речовини, при якій здійснюється круговорот біогенних речовин. У цьому полягає екологічна специфічність функціонування водних екосистем (Леонов, 1991). Окремі внутріводоемнимі процеси впливають на потоки речовини у водному екосистемі і на трансформацію органогенних елементів. До таких процесу відносяться взаємодії і обмін речовиною на кордонах вода-дно і вода-повітря, перерозподіл речовини по вертикалі і горизонталі в межах водойми і при взаємодії з іншими об'єктами, надходження речовини з локальних і розосереджених джерел. Всі ці процеси необхідно кількісно оцінити. Це може бути виконано при вивченні трансформації речовини шляхом розгляду моделей структурно взаємопов'язаних компонентів біогенних елементів (окремо для кожного з них або спільно). Аналіз умов функціонування таких систем дозволяє вивчати специфіку розвитку біотрансформації речовин, роль компонентів водних екосистем і внутріводоемнимі процесів. За допомогою імітаційного математичного моделювання представляється можливим об'єднання різноманітної інформації про стан тих чи інших водних екосистем та отримання комплексної оцінки розвитку процесів трансформації речовин різного походження та природи, відтворення внутрішньорічної і міжрічної динаміки розчинених і зважених компонентів, оцінка впливу окремих внутріводоемнимі процесів на характер просторово-часової мінливості концентрацій речовин і розподілу компонентів екосистеми.
Трансформація речовини і енергії у водних екосистемах здійснюється всередині трофічних мереж. У багатьох спільнотах трофічні мережі включають велику кількість всеїдних організмів, що в переважній більшості випадків вкрай ускладнює або робить просто неможливим віднесення конкретних представників до певних трофічних рівнів (Алімов, 2001; Patten, 1985). Крім того, трофічні зв'язки характеризуються наявністю "петель" замкнутих циклів, що накладаються на основну послідовність цих зв'язків. Найбільш яскраво такі ситуації представлені в "мікробіальних петлях". Все це дозволяє ставити під сумнів існування дискретних трофічних рівнів. Однак це не скасовує справедливості виділення генеральної трофічної послідовності в екосистемах, названої трофічними рівнями. У деяких випадках і для опису найбільш загальних структурних і функціональних закономірностей в екосистемах використання поняття трофічних рівнів виправдано і необхідно.
За відсутності екстремальних умов чисельність популяцій і співтовариств організмів регулюється або обмеженням ресурсів, або пресом хижаків. Такі шляхи регуляції відповідно до теорій регуляції і трофічного каскаду відносяться до контролю "зверху" (top-down) і "знизу" (bottom-up). При цьому, як це випливає з теорії трофічних рівнів, потік біогенних елементів проходить по лінійної вертикальної харчового ланцюга, і рециклінг їх відбувається уздовж цієї ж ланцюга (Бізін, 1997; Carpenter et al., 1987).
Структурна і функціональна стійкість екосистем забезпечується за рахунок складних регуляторних процесів, серед яких велике значення мають позитивні і негативні зворотні зв'язки (Stone, Weisburd, 1992). Негативні зворотні зв'язки виступають як самокорегуюча. Позитивні зворотні зв'язки можуть відігравати стабілізуючу роль. Такий фундаментальний процес в екосистемах як рециклинг біогенних елементів в простій формі може бути представлений у вигляді позитивного зворотного зв'язку. Такий зв'язок здійснюється в так званих "мікробіальних петлях", які можуть розглядатися як справжні "пастки биогенов". Завдяки великій площі мікроорганізмів по відношенню до власної масі і малої швидкості їх занурення створюються умови для швидкого рециклінгу биогенов в поверхневих шарах води і їх повернення в харчовий ланцюг. Ці процеси здійснюються шляхом утилізації мікроорганізмами прижиттєвих виділень фітопланктону, зоопланктону, гідролізу органічних частинок і т. Д. У результаті високої активності мікроорганізмів біогенні елементи багаторазово використовуються фітопланктоном. Вони, будучи замкнутими в "мікробіальних петлях", одночасно створюють додатковий резервуар биогенов. Біомаса мікроорганізмів зростає з високими швидкостями і це призводить до виникнення потужної петлі позитивного зворотного зв'язку в самій основі трофічної мережі, здатної стати своєрідним буфером на шляху трофічного каскаду (Бізін, 1997). Через ці петлі в екосистемах, особливо в пелагічних їх частинах, можуть йти основні потоки речовини і енергії. Так, наприклад, в деяких водоймах через мікробіальних петлі проходить до 50-60% від чистої продукції планктону (Бізін, 1997; Hairston, Hairston, 1993).
Дуже мала частка чистої первинної продукції макрофітів споживається безпосередньо тваринами фитофагами. В основному вона утилізується спільнотами мікроорганізмів. Тому продукція автотрофов, створювана в літоральної зоні, не потрапляє в пелагическую харчовий ланцюг. Обмін енергією між літоральної і пелагическими зонами може здійснюватися за рахунок мобільності організмів вищих трофічних рівнів, наприклад, риб. Однак поки розрахунки такого роду не проводилися.
"Буферний" характер реакції природних екосистем на зовнішні впливи означає, що існує рівень впливу, який можна назвати пороговим, або критичним (Базикін, 1985). При відносно слабких допорогових впливах система як би поглинає, адсорбує впливу і зазнає лише слабкі, кількісні, часто недоступні для спостерігача зміни. Це властивість природних екологічних систем до певної межі протистояти зовнішнім впливам отримало назву "еластичності" екологічних систем (Holling, 1973). Після того як інтенсивність зовнішнього впливу перевищує критичний рівень, система не витримує зовнішнього тиску, ламається і переходить в якісно інше, як правило, небажаний стан. Важливою особливістю такого якісного переходу є його практична незворотність. Зняття впливу не призводить до відновлення колишнього стану екосистеми. Його також неможливо штучно синтезувати. Воно може відтворитися лише в процесі екологічної сукцесії, що вимагає десятків і сотень років.
Контрінтуітівное характер реакції природних екосистем на зовнішні впливи означає суперечить "здоровому глузду" реакцію складної системи на них. У природних системах подібний характер реакції швидше правило, ніж виняток (Базикін, 1985). Яскравим прикладом такої поведінки може служити явище евтрофікації прісноводних водойм. Надходження в озера і водосховища мінеральних добрив часто не тільки не підвищує корисну продуктивність водойм, як можна було б очікувати, а докорінно перебудовує всю структуру екологічної системи і веде її до екологічної загибелі.
В даний час співіснують дві основні точки зору на трофічну структуру водних екосистем: класична, відома як концепція лінійної харчового ланцюга, і її модифікація, яка розглядає мікробіальних спільноти як окрему трофічну щабель. Друга точка зору відома як концепція микробиальной трофічної мережі. Вона постулює активну участь мікроорганізмів (бактерій і найпростіших) в потоці речовини і енергії (Бульйон, 2002).
Велике значення мікроорганізмів як проміжних ланок в перенесенні енергії "первопіщі" до верхніх трофічних рівнів вперше виявлено і сформульовано для морських екосистем (Сорокін, 1971; Сорокін, 1973). "Первопіща" (термін, введений Ю. І. Сорокіним (Сорокін, 1973)) об'єднує в себе первинну продукцію і "мертве" органічна речовина (детрит і розчинені органічні речовини). За рахунок її утилізації ступінь участі бактерій і найпростіших в метаболізмі гетеротрофною частини планктонного спільноти досягає 80 - 90%.
Слід підкреслити неподільність концепцій лінійної харчового ланцюга і микробиальной трофічної мережі, так як друга концепція випливає безпосередньо з першої, тобто микробиальная трофічна мережа виконує функцію шунта в харчовому ланцюгу, коли продукція фітопланктону по тій або іншій причині (наприклад, через домінування колоніальних водоростей ) не може бути прямо використана планктонними тваринами. У гідроекології поряд з термінами "микробиальная трофічна ланцюг" і "мікробіальних співтовариство" часто використовується термін "микробиальная петля".
Розділ включає такі підрозділи::
Додаткова інформація:
© 2001-2019 Кафедра біофізики МГУ
При моделюванні таких систем необхідно завжди задаватися питанням, "про які масштабах часу йде мова?