
VIVOS VOCO: Н.С. Жданова, "Нитка Аріадни в генетиці"
Нитка Аріадни в генетиці
Н.С. Жданова
Наталія Сергіївна Жданова, д.б.н., вед. наук. співр. лаб. генетики розвитку.
Карти геномів тварин і рослин - це нитка Аріадни, дотримуючись якої, генетики шукають потрібні гени, вивчають, як організований геном і як він еволюціонує, як успадковуються гени і ознаки. Ця нитка веде дослідників і в світ суто практичних турбот - допомагає зрозуміти, як в нормальному і хворому організмах взаємодіють гени та їх продукти, відшукати оптимальні стратегії селекції сільськогосподарський тварин і організувати медико-генетичне консультування. Особливо необхідні докладні карти геномів людини, а також тих видів ссавців, що використовуються в якості модельних об'єктів в генетичних дослідженнях.
На службі міжвидові клітинні гібриди
В даний час геномні карти представляють собою результат об'єднання генетичних, цитогенетичних і фізичних карт, а також даних про секвенування ДНК за все генома або окремих хромосомних районів. Така карта виглядає як схема, на якій в певному порядку і на певній відстані один від одного нанесені маркери, роль яких можуть грати або морфологічні і фізіологічні ознаки організму, або гени та їх продукти, або окремі послідовності ДНК.
Різні типи карт відрізняються способом отримання. На початку XX ст. картування проводилося на основі гибридологического аналізу: вивчалося успадкування ознак в першому і другому поколіннях гібридів від схрещування особин, що розрізняються за кількома ознаками. Однак через тривалість репродуктивного періоду, малого числа нащадків і відсутності в той час методів виявлення поліморфних ознак докладні генетичні карти ссавців створені лише недавно.
Прорив в картуванні генома людини та інших видів ссавців настав в 60-х роках минулого століття [ 1 ]. Тоді з'явилася ідея замінити на схрещування осіб - злиття їх генеративних клітин - злиттям соматичних клітин різних видів тварин. Цьому передували багаторічні дослідження по необмеженому розмноженню клітин поза організмом (in vitro) в спеціальних середовищах на склі або пластикових підкладках. Такі клітинні культури отримали назву тривало перевіваемих. Клітини цих ліній за деякими властивостями відрізняються від звичайних, так як більшість веде своє походження від пухлинних клітин, що містять перебудовані хромосоми.
Щоб клітинні гібриди могли необмежено розмножуватися in vitro, необхідно в якості хоча б одного з батьків використовувати саме тривало перещеплюваних клітини. Роль другого з батьків зазвичай грають нормальні клітини, виділені безпосередньо з організму. Для цієї мети зручні лейкоцити периферичної крові. При спонтанному або індукованому злиття клітин спочатку формуються багатоядерні клітини (гомо- або гетерокаріони), а потім, коли зіллються ядра, утворюється гібридна клітина. Розмножилися, вона дає гібридний клон.
Довго вважалося, що спонтанно зливаються клітини тільки в культурах, а в організмі, якщо це і трапляється, відбір вибраковує виникли гібриди. Однак недавно дослідники спостерігали спонтанне злиття трансплантованих клітин кісткового мозку з мутантними гепатоцитами мишей. В результаті утворювалися гібридні клітини, в яких відновлювалася втрачена раніше функція гепатоцитів.
Спочатку область застосування міжвидових гібридів соматичних клітин була досить великою. Тривалий час на гетерокаріонов і соматичних гібридах вивчалися реактивация геномів, активація і придушення експресії генів, роль в цих процесах ядра і цитоплазми. Використовуючи клітинні гібриди, вперше вдалося показати, що в нормальних клітинах людини існують гени, що пригнічують злоякісний ріст. У теперішній же час такі гібриди використовуються в основному для картування хромосом. Як же за допомогою гібридів картировать гени?
Уже в перших роботах по вивченню хромосомного складу міжвидових клітинних гібридів з'ясувалося, що вони випадковим чином втрачають хромосоми одного з батьківських видів (як правило, батьків з нормальним каріотипом). Це властивість привернуло творців генетичних карт, оскільки з втратою хромосоми в гібриді зникають і локалізовані в ній маркери.
Першу спробу пов'язати локалізацію гена зі збереженням в гібридах "людина-миша" певної хромосоми людини зробили на початку 70-х років. Оскільки в гібридних клонах була виявлена кореляція між наявністю в них хромосоми 17 людини і тимідинкінази 1, то ген ферменту був віднесений саме до цієї хромосомі (рис.1).
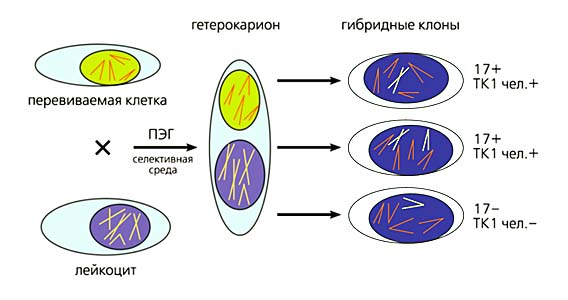
Мал. 1. Схема отримання гібридних клонів "людина-миша" і локалізації гена тимідинкінази 1 (ТК1).
Тривало перещеплюваних клітини миші, в яких цього гена немає, і лейкоцити периферичної крові людини обробляють поліетиленгліколем (ПЕГ), в результаті чого структура мембран порушується так, що ті і інші клітини злипаються, а потім зливаються, утворюючи дво- або багатоядерні гетерокаріони.Після синхронізації і вступу ядер в мітоз або за рахунок їх прямого злиття формується гібридна клітина з ядром, що містить хромосоми обох батьківських видів.У використовуваної селективної середовищі гинуть мишачі клітини, що не мають активного гена ТК1 (необхідного для біосинтезу ДНК), і лейкоцити людини, оскільки без спеціальних стимуляторів вони in vitro не діляться.Виживають лише гібридні клітини, в які лейкоцити привнесли ген ТК1.З таких клітин утворюються гібридні клони, і з них створюють статистичну Картира панель.Через випадкових втрат людських хромосом різні клони містять різні хромосоми.Тут показані дві клітини клонів з хромосомою 17 (хр. 17 +) людини, а оскільки в них проявляється активність ТК1 (+), її ген був віднесений саме до цієї хромосомі.
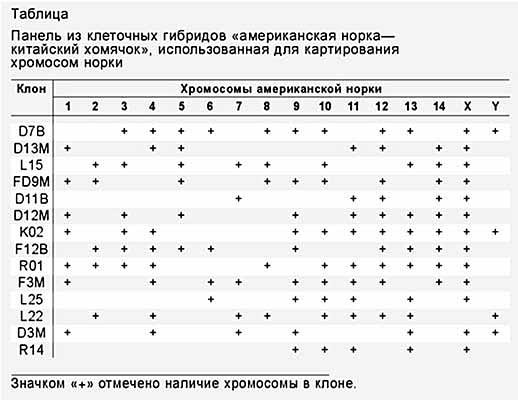
В середині 80-х років основним інструментом картування хромосом людини стали панелі гібридних клонів. Панель - це набір клітинних гібридів, в яких статистично представлений весь геном досліджуваного виду (таблиця). Як правило, за допомогою клітинних гібридів можна визначити, в якій саме хромосомі локалізовано ті чи інші маркери. Якщо вони знаходяться в одній і тій же хромосомі, їх називають "сінтеннимі". Однак цим способом можна картировать тільки ті ознаки, які виявляють міжвидової поліморфізм, тобто відрізняються у батьківських видів з яких-небудь параметрами. Крім того, картіруемий ознака не повинен придушуватися або активуватися в гібридах. Уже в 80-х роках на карту хромосом людини було нанесено близько 600 генів. Ці успіхи відкрили дорогу для картування хромосом інших видів ссавців.
З використанням панелі гібридів клітин американської норки і бурозубки звичайної з клітинами миші або китайського хом'ячка в нашому інституті побудовані карти хромосом норки і бурозубки. У хромосомах бурозубки було локалізовано 49 генів і чотири високо поліморфних просто виявляються маркера - мікросателіта * (рис.2), а в хромосомах норки - близько 130 генів і 40 мікросателітів [ 2 ]. І хоча останні не кодують жодних ознак, а функція їх остаточно не ясна, вони цілком придатні як реперів на карті, до яких можна "прив'язати" важливі для нас ознаки. Вивчивши, наприклад, зчеплення між мікросателіти і генами забарвлення американської норки, вдасться локалізувати ці гени в хромосомах і використовувати мікросателіти для оптимізації селекції норок з потрібною забарвленням хутра.
* Мікросателіти - короткі послідовності ДНК, в яких мотив з декількох нуклеотидів повторений багато разів. 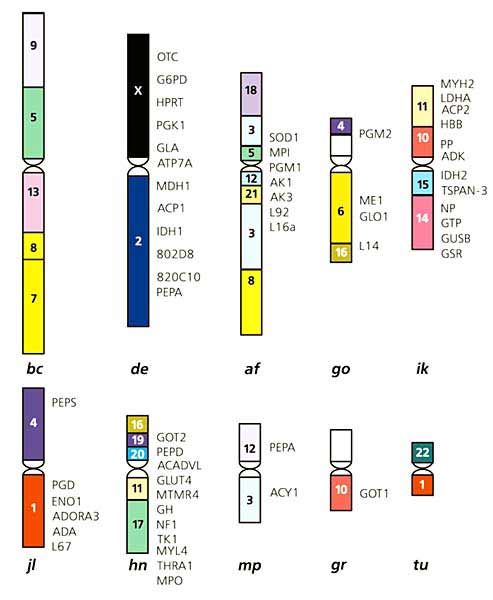
Мал. 2. Сінтенная карта хромосом бурозубки звичайної.
Під кожною схематично зображеної хромосомою приведено її позначення (латинські літери),
а праворуч - гени і мікросателіти (буквено-чисельні символи);
кольором відзначені консервативні райони хромосом людини (цифри і числа) в хромосомах бурозубки.
Ідентифікація в клітинних гібридах всього генетичного матеріалу виду необхідна для використання їх в картуванні. Чи не виявлені хромосомні перебудови стануть джерелом помилок, але якщо перебудови ідентифіковані, вони послужать для внутріхромосомной локалізації маркерів, тобто віднесення їх до певного хромосомному району. Подібний приклад - клітинні гібриди свині. Вони містили велику кількість перебудованих хромосом, що надовго затримало їх картування. Але коли в гібридних клонах методами молекулярної цитогенетики були ідентифіковані фрагменти, з яких складалися перебудовані хромосоми, з'явилася можливість віднести маркери до конкретних хромосомним районам (рис.3, 4).

Мал.3.Результати ідентифікації дев'ятої хромосоми свині в гібридному клон "свиня-американська норка".Зліва: ідентифіковані диференціальної забарвленням барвником Гімза хромосоми свині на тлі хромосом норки, де стрілкою вказано положення дев'ятої хромосоми.В середині: вона ж (вказана стрілкою) в препараті, отриманому в результаті прямої гібридизації in situ (ДНК свині була гібрідізоваться з хромосомами клону).Справа: чітко видимі дві світлі постаті - це хромосоми свині, виявлені після зворотної гібридизації (фракція ДНК свині з клону гібрідізоваться з її хромосомами).
Мал.4. Мікрофотографії диференційно забарвлених нормальної (N) другий хромосоми норки і трьох перебудованих варіантів (стрілкою вказана транслокация фрагмента хромосоми 2 на хромосому хом'ячка) та схема хромосоми з нанесеними на неї генами.Для картування служили перебудовані хромосоми з різних гібридних клонів "американська норка-китайський хом'ячок".На схемі дужками вказані райони локалізації картірованних генів, а їх позначення дано символами;p і q - плечі хромосоми.радіаційні гібриди
Якщо перед злиттям клітин спеціально розірвати хромосоми, чи можна використовувати отримані таким чином гібриди для картування? Виявилося, можна. Йдеться про особливий тип міжвидових клітинних гібридів - радіаційних. Вони утворюються в результаті злиття перевіваемих клітин і нормальних, які попередньо опромінюють летальною дозою радіації [ 3 ].
У радіаційних гібридах статистично зберігаються не тільки фрагменти хромосом, що містять центромеру і гени, необхідні для виживання клонів, а й фрагменти, перенесені на партнерські хромосоми (рис.5). Вдалося навіть отримати радіаційні гібриди між клітинами ссавців і риб та використовувати для картування хромосом останніх.
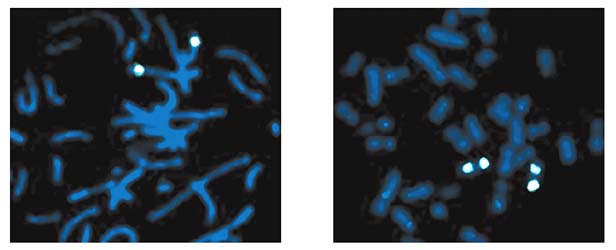
Мал.5. Результати ідентифікації генетичного матеріалу свині в радіаційному гібриді, отриманому від злиття перевіваемих клітин китайського хом'ячка і клітин свині, опромінених дозою 6000 радий.Прямий гибридизацией виявлені в цьому клоні два фрагмента (світлі постаті) хромосоми свині, перенесені на хромосоми хом'ячка (зліва), а зворотної гибридизацией отримано доказ, що ці фрагменти є райони другий хромосоми свині.
В основі радіаційного картування геномів лежать роботи англійських дослідників С.Госса і Г.Харріса [ 4 - 6 ]. Вони не тільки випередили час на чверть століття, але і окреслили більшість напрямків у вивченні радіаційних гібридів як феномена і в використанні їх для картування хромосом. Спосіб отримання таких гібридів до сих пір залишився незмінним, змінилися лише методи статистичної обробки експериментальних даних. Створені комп'ютерні програми дозволяють одночасно аналізувати кілька тисяч маркерів.
Залежно від дози опромінення нормальних клітин виходять панелі, завдяки яким можна побудувати радіаційні карти різного ступеня дозволу. Спочатку були отримані радіаційні гібриди від злиття з клітинами, опроміненими дозою в діапазоні 3-10 тис. Радий. Використовуючи панелі людини з таких гібридів, американські вчені визначили порядок близько 6 тис. Мікросателітів і маркерів експресуються генів. І хоча в порівнянні з іншими картами людини ці були більш високої роздільної здатності, на них маркери на відстані 4 тис. Пар нуклеотидів поділялися з невисоким ступенем достовірності. Щоб підвищити дозвіл радіаційних карт, дослідники отримали панелі, що складаються з гібридів з нормальними клітинами людини, опроміненими більш високою дозою радіації, 40 тис. Радий. На сконструйованої карті з високим ступенем достовірності поділялися маркери на відстані всього в 100 пар нуклеотидів. Ціна високого ступеня дозволу - неможливість вибудувати протяжні ділянки карти. Як правило, таке картування служить для вивчення генетичного оточення цікавлять локусів. Воно застосоване, наприклад, при дослідженні хромосомних районів свині, в яких локалізовані головні гени, що впливають на якість м'яса або число нащадків в посліді. Таким чином, за допомогою радіаційного картування в даний час отримують найдокладніші хромосомні карти.
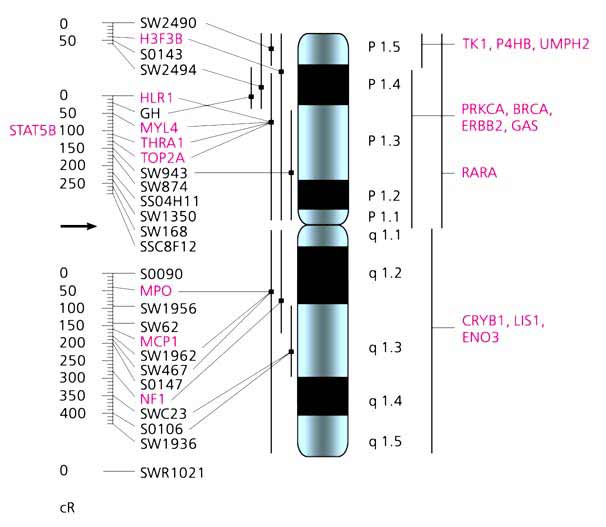
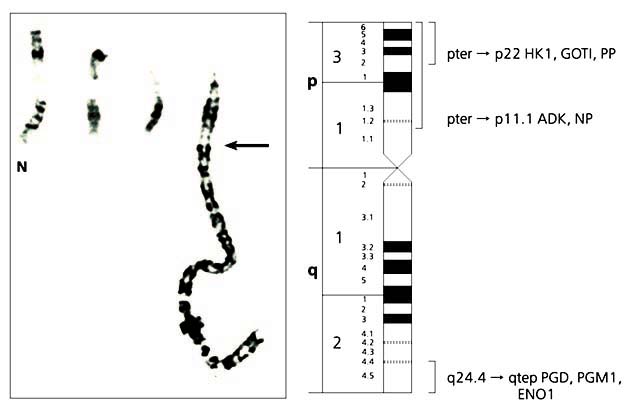
Мал. 6. Радіаційна (зліва) і цитогенетична карти хромосоми 12 свині.
Карта побудована за допомогою радіаційної панелі, що містить 118 клонів, отриманих французькими вченими в результаті злиття клітин свині, опромінених 7000 радий, з клітинами китайського хом'ячка [7].Зразки ДНК панелі були передані вченим різних країн, що дозволило швидко побудувати радіаційну карту.Картірованниє нами гени позначені червоним шрифтом, а мікросателіти, картірованниє іншими вченими, - чорним.Вертикальними лініями уздовж цитогенетичної карти диференційно пофарбованої хромосоми позначені райони локалізації деяких маркерів в хромосомі.Відносні відстані між маркерами на радіаційної карті дані в умовних одиницях - cR;стрілкою вказано можливе положення центромери.Отже, на основі радіаційної панелі вдається вибудувати в певному порядку сотні і тисячі маркерів і визначити між ними відносні відстані. На виході отримують якесь число сінтенних груп маркерів, зазвичай перевищує кількість хромосом у виду. Іншими словами, група зчеплення однієї хромосоми виявляється роздробленою. Точки розривів майже завжди розташовані в центромерних і субтеломерних районах хромосом, але можуть перебувати і в інших місцях. Розірваність карти в першу чергу відображає нерівномірність розподілу радіаційних розривів по довжині хромосоми. Як "прив'язати" сінтенние групи до конкретних хромосомами і орієнтувати по їх довжині? Для цього кілька маркерів з кожної групи необхідно локалізувати безпосередньо в хромосомах за допомогою цитогенетичних методів картування, наприклад флуоресцентної гибридизацией in situ, яка дозволяє побачити, в якій хромосомі і в якому її районі знаходиться маркер (рис.6).
Все пізнається в порівнянні
У перших же роботах з картування геномів ссавців за допомогою міжвидових гібридів соматичних клітин з'ясувалося, що гени, сінтенние у одного виду, сінтенни і у інших видів. Дивно, але деякі асоціації генів простежуються аж до птахів і костистих риб. Наявність в геномах таких консервативних, або гомеологічних, районів викликало великий інтерес у дослідників, і в подальшому сформувався цілий напрямок - порівняльне картування, яке увійшло до складу геноміки, науки про структуру і еволюції геномів.
Консервативними бувають і цілі хромосоми. Наприклад, один і той же набір генів містять хромосома 17, 19-а корови, 12-я свині і т.д. Інші консервативні ділянки істотно менше за розміром. Порівняльне картування недарма стало самостійною гілкою геноміки - воно дає масу цікавої інформації. Так, зіставленням радіаційних карт ссавців з результатами секвенування геномів людини і миші встановлено, що в сайтах еволюційних розривів між великими консервативними районами, як правило, локалізовані буквально одиничні гени або невеликі послідовності розміром близько мільйона пар нуклеотидів з інших, теж консервативних, районів. Цікавими в цьому відношенні виявилися періцентромерние області хромосом (рис.7). У свині, наприклад, в них містяться маркери, які не належать власним консервативним ділянкам періцентромери. Це свідчить про високу швидкість хромосомних перебудов в тих місцях, де в ході еволюції кариотипов утворюються нові центромери.
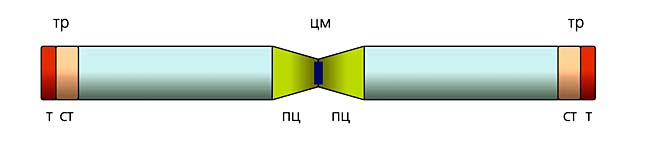
Мал.7. Основні елементи структури хромосом.Центромера (ЦМ) - це нуклеопротеїдні комплекс, что складається з відоспеціфічності повторення послідовностей ДНК и однаково у всех ссавців білків, які відповідають за правильну поведение хромосом во время ділення клітін.Періцентромера (ПЦ) прілягає безпосередно до центромере и збагачена різного роду, что повторюються.Термінальна (ТР) частина хромосом Включає теломери (Т) и субтеломеру (СТ).Теломера у всех ссавців складається з багаторазове тандемно повторення мотиву з шести нуклеотидів.Ее спеціфічна організація и Механізм реплікації ДНК оберігають хромосоми від злиттів и обумовлюють їх цілісність.Субтеломера - це буфер между теломер и Основним тілом хромосоми, де локалізовані гени, и містіть велику Кількість повторень ділянок ДНК.У субтеломерах хромосом людини є деградовані теломерні повтори, інші подвоєні послідовності і сегментні дуплікації;у інших видів ссавців тут може міститися сателітна ДНК - короткий мотив, повторений до 100 тис. разів.
За допомогою порівняльного картування вдалося показати, що швидкості, з якими еволюціонували каріотипи різних видів ссавців, істотно різняться. Так, предкові геноми плацентарних тварин і мишоподібних гризунів відділені, мабуть, більш ніж 150-ю хромосомними перебудовами, а хижих - тільки дев'ятьма. Геном свині утворився за рахунок 42 перебудов предкового генома парнокопитних, а геном корови - за рахунок 99. Фактично в кожному загоні або сімействі описані і повільно, і швидко еволюціонують види. Людина, шимпанзе і гібон - хоча і родичі, але відносяться до різних родин приматів. Однак каріотипи людини і шимпанзе ідентичні, за винятком злиття декількох хромосом, тоді як 26 хромосом гібон складені з 70 фрагментів хромосом людини. Таким чином, швидкість реорганізації кариотипов різних видів не завжди пов'язана з їх еволюційним положенням.
Судячи з радіаційним картками, в геномах риби Danio rerio і людини як мінімум 247 гомологічних сегментів. І ця величина порівнянна з тією (201), яка на тому ж рівні дозволу карт була визначена для миші і людини. Виявилося, що кожна хромосома D.rerio містить від двох до семи блоків, що зустрічаються в різних хромосомах людини. Ще приклади невідповідності: число консервативних районів в геномах курки і людини становить 154, а курки і миші - 312.
Складається враження, що геноми, принаймні ссавців, складаються в основному з блоків, які у різних видів з'єднані в різних комбінаціях. Обмеження накладені, очевидно, на варіанти кариотипов, "незручних" для протікання таких головних клітинних процесів, як мітоз і мейоз, або порушують допустиму архітектоніку ядра.
При порівнянні радіаційних карт виявлена нова еволюційна перебудова - зміна положення центромери в консервативних районах. Цей феномен описаний майже для всіх хромосом корови. Такий же тип перебудови знайшли і ми, зіставивши радіаційні карти декількох хромосом свині і людини. Незважаючи на те, що центромера відповідає за дуже важливу функцію - правильна поведінка хромосом в мітозі і мейозі, - її положення в консервативних районах виявилося вельми лабільним.
Таким чином, порівняння радіаційних карт різних видів дозволяє виявляти склад, межі та порядок генів в консервативних районах, реконструювати їх предковий склад і предкові каріотипи, визначати швидкість реорганізації кариотипов в різних гілках еволюції.
Наявність консервативних районів мимоволі викликає питання: де рвалися хромосоми, коли формувався новий каріотип, в випадкових або певних місцях? Довгий час відповідали - "в випадкових". Але в останні роки стали з'являтися дані про те, що це абсолютно певні хромосомні сайти. Більш того, в еволюції кариотипов ссавців одні й ті ж райони неодноразово використовувалися в якості і місць розривів, і місць злиття. Розрізнені факти такого роду постійно звертали на себе увагу дослідників, але закономірністю стали сприйматися після порівняння геномів восьми видів з п'яти загонів ссавців. У числі цих видів - людина, миша і щур з уже повністю розшифрованими геномами, собака, свиня, корова, кішка і коня з радіаційними картами високого дозволу. У каріотипі згаданих тварин виявлено близько 1200 ділянок гомологичной сінтеніі (консервативних районів, в яких збережений не тільки склад, але і порядок генів хоча б у двох видів). Крім того, ідентифіковано 367 хромосомних областей, які поділяли ділянки гомологичной сінтеніі, саме в них і були локалізовані еволюційні розриви. Але найцікавішим виявилося те, що 20% таких районів неодноразово брали участь в реорганізації кариотипов. Решта були специфічними для окремих загонів і видів [ 8 ].
Що ж собою являють ті ділянки хромосом, які неодноразово використовувалися в еволюції кариотипов ссавців? Найчастіше це центромерного, періцентромерние і термінальні області, насичені різного роду повтореними послідовностями. Геноми людини і миші часто містять в таких місцях дупліціроваться гени, причому у миші члени одного і того ж генного сімейства розселені по різних хромосомах (це врешті-решт може призвести до утворення нових генів). У тих же районах знаходяться скупчення L1- і інших некодуючих повторів.
У приматів свій шлях еволюції хромосом?
У геномі людини більше половини ділянок, в яких неодноразово відбувалися еволюційні розриви, містять особливі структури, названі сегментними дуплікації. Вони являють собою багато разів повторені протяжні послідовності, цілі гени або їх частини, і можуть досягати розміру в кілька сотень тисяч пар нуклеотидів. У геномі людини такі послідовності складають 5%. Велика кількість цих структур (близько половини їх утворилося відносно недавно) у приматів - характерна особливість їх геномів. У інших видів ссавців сегментних дуплікацій істотно менше.
В цілому в геномах приматів положення приблизно 90% районів, в яких відбувалися еволюційні розриви, збігається з локалізацією сегментних дуплікацій. Частота порушення сінтеніі в містять їх сайтах в три рази вище, ніж в сайтах, де вони не виявлені. Цей факт, а також наявність таких дуплікацій і в місцях ненарушенной предковой сінтеніі свідчать про те, що вони сприяли появі хромосомних перебудов в геномах приматів, а не були їх наслідком. Таким чином, еволюція їх кариотипов виявилася безпосередньо пов'язаної з особливостями молекулярної еволюції їх геномів.
У людини половина сегментних дуплікацій знаходиться в термінальних (субтеломерних) районах хромосом. Прикладом тому можуть бути дуплікації, в яких зосереджені члени сімейства генів, що кодують нюхові рецептори. Блоки, які містять по три гена з цього сімейства, виявлені в субтеломерах чотирнадцяти хромосом, причому в чотирьох з них ці асоціації утворилися в результаті як мінімум двох обмінів між кінцями хромосом.
З результатів молекулярного аналізу сегментних дуплікацій слід, що більшість з них утворилося за рахунок повторних актів негомологичной і неалельних гомологичной рекомбінації ДНК. Субтеломери - самі швидко еволюціонує райони в геномі людини. У них висока швидкість нуклеотидних замін і дуплікацій не тільки великих, а й невеликих ділянок. За влучним висловом Е.В.Лінардополу, що вивчала субтеломери, вони представляють собою і звалище покидьків молекулярної еволюції геному, і місце, де утворюються нові гени [9]. Така організація пояснює схильність термінальних районів хромосом людини до перебудов.
* * *
Структура геному кожного виду, безсумнівно, формується завдяки тривалому відбору "відповідних" варіантів, і зовсім не байдуже, в якому місці геному знаходиться той чи інший ген. У численних дослідженнях показано, що по довжині хромосом гени розподілені не випадковим чином. Їх щільність вище в районах, збагачених певними нуклеотидами. Як правило, гени, локалізовані в безпосередній близькості один від одного, еволюціонують з подібною швидкістю і об'єднані в еволюційні одиниці, а ті, що експресуються в одній тканини і на конкретній стадії розвитку організму, утворюють в хромосомах щось, що нагадує кластери.
За 30 років, протягом яких активно ведуться роботи по хромосомному картування ссавців, розроблені методи, що дозволяють з високою точністю визначати місце розташування в хромосомах генів, ознак і нуклеотиднихпослідовностей. Отримано геномна карта людини. Наскільки вона докладна, можна судити хоча б по тому, що на неї нанесені майже всі відомі гени. Ця карта зіграла важливу роль при розшифровці людського генома: допомогла вибудувати в певному порядку короткі фрагменти ДНК і перевірити правильність результатів секвенування. З ними разом карту геному можна розглядати як єдину віртуальну карту.
На жаль, самі високоразрешающем фізичні карти - радіаційні - створені лише для десятка видів ссавців. Серед них немає навіть представників усіх загонів цього класу тварин. Побудувати такі карти для більшого числа видів і вивчити, що ж лежить в основі еволюції кариотипов ссавців всіх загонів, а не тільки приматів, - завдання майбутніх досліджень. Образно кажучи, нитка Аріадни, яка веде по лабіринтах генетики, треба ще плести і плести.
література
1. Ефруссі Б. Гібридизація соматичних клітин. М., 1976.
2. Kuznetsov SB, Matveeva NM, Murphy WJ et al. // Journal of Heredity. 2003. V.94. №5. P.386-391.
3. Жданова Н.С. // Генетика. 2002. Т.38. С.581-595.
4. Goss SJ, Harris H. // Nature. 1975. V.255. P.680-684.
5. Goss SJ, Harris H. // J. Cell Sci. 1977. V.215. P.17-37.
6. Goss SJ, Harris H. // J. Cell Sci. 1977. V.215. P.39-57.
7. Yerle M., Pinton P., Robic A. et al. // Cytogenet. Cell Genet. 1998. V.82. P.182-188.
8. Murphy WJ, Larkin DM, Everets-van der Wind A. et al. // Science. 2005. V.309. P.613-617.
9. Linardopoulou EV, Williams EM, Fan Y. et al. // Nature. 2005. V.437. P.94-100.

Як "прив'язати" сінтенние групи до конкретних хромосомами і орієнтувати по їх довжині?
Наявність консервативних районів мимоволі викликає питання: де рвалися хромосоми, коли формувався новий каріотип, в випадкових або певних місцях?
Що ж собою являють ті ділянки хромосом, які неодноразово використовувалися в еволюції кариотипов ссавців?
У приматів свій шлях еволюції хромосом?